Honda Group, Colombia
| Honda Group Stratigraphic range: Late Oligocene-Late Miocene typically Middle Miocene (Laventan) ~13.8–11.8 Ma PreЄ Є O S D C P T J K Pg N ↓ | |
|---|---|
 Statue of a mammal from the Honda Group in Villavieja, Huila | |
| Type | Geological group |
| Sub-units | Villavieja Fm. Cerro Colorado Mb. Baraya Mb. La Victoria Fm. Cerbatana Mb. |
| Underlies | Gigante Group Neiva Fm. |
| Overlies | Payandé Group Barzalosa Fm., Saldaña Fm., Santa Teresa Fm. |
| Thickness | up to 5,000 m (16,000 ft) |
| Lithology | |
| Primary | Conglomerate, sandstone |
| Other | Claystone, siltstone, volcanoclastic sediments |
| Location | |
| Coordinates | 5°11′31″N 74°43′21″W / 5.19194°N 74.72250°W / 5.19194; -74.72250Coordinates: 5°11′31″N 74°43′21″W / 5.19194°N 74.72250°W / 5.19194; -74.72250 |
| Region | Huila, Tolima, Cundinamarca Middle Magdalena Basin, Upper Magdalena Basin Central & Eastern Ranges Andes |
| Country | |
| Extent | ~250 km (160 mi) |
| Type section | |
| Named for | Honda |
| Named by | Hettner |
| Location | Honda (original) Tatacoa Desert (redefined) |
| Year defined | 1892 |
| Coordinates | 3°14′04″N 75°12′15″W / 3.23444°N 75.20417°W / 3.23444; -75.20417 |
| Approximate paleocoordinates | 3°06′N 72°24′W / 3.1°N 72.4°W / 3.1; -72.4 |
| Region | Tolima (original) Huila (redefined) |
| Country | |
| Thickness at type section | 3,255 m (10,679 ft) |
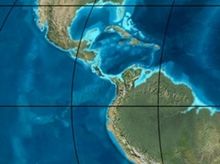 Paleogeography of Northern South America 20 Ma, by Ron Blakey | |

The Honda Group (Spanish: Grupo Honda, Tsh, Ngh) is a geological group of the Upper and Middle Magdalena Basins and the adjacent Central and Eastern Ranges of the Colombian Andes. The group, in older literature also defined as formation, is in its present-day type section in the Tatacoa Desert in the department of Huila subdivided into two main formations; La Victoria and Villavieja.
The group was originally defined in and named after Honda, Tolima, but has been redefined based on the many fossil finds in the Tatacoa Desert, 250 kilometres (160 mi) to the south. In the original type section of its occurrence, the 3,255 metres (10,679 ft) thick group is subdivided into three formations, from old to young; Cambrás, San Antonio and Los Limones.
The group dates to the Neogene period; in its broadest definition from the Late Oligocene to Late Miocene, and in the redefined type section restricted to the Laventan age of the South American Land Mammal Ages (SALMA), equivalent to the Middle Miocene Serravallian epoch.
The Honda Group is a Konzentrat-Lagerstätte at the fossiliferous La Venta site in the department of Huila and eastern Tolima and hosts one of the richest formations containing Miocene fauna worldwide.
Contents
1 Etymology and definitions
2 Regional setting
3 Description
3.1 Stratigraphy
3.1.1 La Victoria Formation
3.1.2 Villavieja Formation
3.1.3 Honda area
3.2 Age
3.3 Depositional environment
3.3.1 Paleoclimate and vegetation
3.4 Petroleum geology
4 Fossil content
4.1 Mammals
4.2 Birds
4.3 Reptiles and amphibians
4.4 Fish and crustaceans
5 Panorama
6 See also
7 References
7.1 Bibliography
7.1.1 Regional geology
7.1.2 Local geology
7.1.3 Paleontology
7.1.4 Maps
8 External links
Etymology and definitions

View of Honda, Tolima, namesake of the Honda Group
The group was first defined by Hettner in 1892 in the area of the town after which it is named; Honda, Tolima.[1] In 1942 and 1946, the group was defined as a formation by Royo and Gómez. The first author who used the name Honda for a group, was American zoologist who studied the La Venta fauna in detail, Ruben Arthur Stirton. Subdivisions of the group have been proposed by many different authors with high detail in the different beds.[2] Names as "Monkey Beds", "Fish Beds" and "Unit above Fish Beds" have been colloquially used to designate certain stratigraphic units based on their fossil content.[3][4] The most accepted definition of the group, formations and members was proposed by Villarroel et al. in 1996 to simplify the stratigraphy in a regional sense solving the excessive subdivision into 19 units by Guerrero et al. (1994).[5]
Regional setting
Today, the sediments of the Honda Group are exposed in the Middle (Valle Medio del Magdalena, VMM) and Upper Magdalena Valley (Valle Superior del Magdalena, VSM) in an intermontane valley between the Central Ranges in the west and the Eastern Ranges in the east. The valley in between the two major orogenic chains is filled by the Magdalena River, the longest river of Colombia. The Upper Magdalena Valley geologically is subdivided into the Neiva Sub-basin with the Girardot Sub-basin of the southernmost Middle Magdalena Valley to the north, divided by the Natagaima Arch.[6] The Neiva Sub-basin is bound by the Chusma Fault in the west and to the east by the west-verging Garzón Fault.[7] The Mulato-Getudo Fault possibly underlies the Honda Group south of the La Miel River.[8] The Tatacoa Desert, where the type section of the Honda Group has been redefined is an unusually dry region in Colombia, caused by a mountain induced rain-shadow effect. In Middle Miocene times, the geography was more comparable to the present eastern foothills of the Andes.[9]
The tectonic history of the three mountain chains of Colombia, from west to east, the Western, Central and Eastern Ranges has been studied in detail. The Western and Central Ranges were the first to be exhumed in the Paleogene, with minor uplifts in the Eastern Ranges at this age. The onset of the regional uplift of the Eastern Ranges is dated around the Middle Miocene, with an increased rate of exhumation between 12 and 3 Ma.[10] This caused a segmentation of the current Magdalena Basins and the Llanos Basin to the east, previously part of a larger foreland basin to the east of the Central Ranges.[11] The León Formation in the Llanos Basin provides the first indication of the tectonic uplift of the Eastern Ranges, isolating the Llanos Basin from the Magdalena Valley.[12]
Description
Stratigraphy
The Honda Group unconformably overlies in parts the volcanic deposits of the Prado Member,[4]Barzalosa Formation of the Payandé Group, and in other parts the Saldaña,[13] and Santa Teresa Formations.[14] The unit is overlain by the Neiva Formation of the Gigante Group.[3][15] The presence of a hiatus between the Honda Group and the Barzalosa Formation has been suggested.[16] The group is characterised by two main formations; the lower La Victoria Formation and upper Villavieja Formation. Previously, the La Dorada Formation has been named as a subdivision of the Honda Group, while other authors define that unit as a member. Other names for members and formations are Cerbatana Member, also published as Cervetana Member, named after Quebrada La Cerbatana, Las Mesitas Formation, El Líbano Formation, Baraya Volcanic Member, named after Baraya, and Cerro Colorado Red Bed Member.[17] The Perico Member of the La Dorada Formation has been made equivalent to the La Victoria Formation, as well as the El Líbano Formation.[4]
La Victoria Formation
This formation was first defined by Guerrero in his Master's thesis (1991) and further refined in his doctoral thesis in 1993. The type locality of the La Victoria Formation has been set in the La Venta area and the formation is named after the town of La Victoria, at 15 kilometres (9.3 mi) north-northeast of the urban centre of Villavieja. The formation, restricted to the Neiva Sub-basin,[18] consists mainly of sandstones, conglomeratic sandstones and conglomerates (75%) with intercalated claystones and siltstones (25%).[5]
The upper part of the La Victoria Formation, underlying the Villavieja Formation, is characterised by a 45 metres (148 ft) section of conglomerates, designated the Cerbatana Member or Cerbatana Conglomerates. The conglomerate shows trough-cross lamination and imbrication of clasts in a predominantly matrix-supported sequence, with minor clast-supported sections. The base of the conglomerate marks an erosional surface into the underlying silt and clay beds. The clasts of the conglomerate are mostly milky quartz, chert and volcanic in origin, with diameters averaging around 7 centimetres (2.8 in) with a maximum of 15 centimetres (5.9 in). Conglomeratic and medium to coarse-grained sandstone banks, with a similar grain composition as the conglomerates, up to 2 metres (79 in) thick are intercalated between the conglomeratic sections. The sandstones are cemented by calcium carbonate in hardgrounds that sometimes form rounded concretions. The claystones and siltstones that are less commonly found in the La Victoria Formations range in thickness from 1 to 11 metres (3.3 to 36.1 ft) and show reddish-brown, greenish-grey and greyish colourations.[19]
Villavieja Formation

Wind erosion of the Villavieja Formation in the Tatacoa Desert
The name Villavieja Formation was first proposed by Wellman in 1968 as a member of the Honda Formation. Two years later, the author elevated the rank to a formation, as part of the Honda Group. The formation takes its name from the municipality Villavieja, Huila, 35 kilometres (22 mi) to the north-northeast of the departmental capital Neiva. The type locality is situated on the right bank of the Magdalena River in the Eastern Ranges of the Colombian Andes.[20]
The contact between the Villavieja Formation and the underlying La Victoria Formation is concordant.[18] The basal part of the Villavieja Formation comprises siltstones and claystones that also form the bulk of the formation with 75 percent. The remaining quarter is composed of conglomeratic sandstones. The thickness of the siltstones and claystones can exceed 8 metres (26 ft) and have interspersed fine- to medium-grained 10 centimetres (3.9 in) thin sandstone beds. The fine sediments of the Villavieja Formation are coloured greenish, reddish-brown or bluish-grey and display weathering patterns in so-called "cauliflower erosion" structures. The light grey coarser beds, up to conglomeratic sandstone size, do not exceed 2.5 metres (8.2 ft) in thickness and commonly show a lateral transition with the silt- and claystones. The Baraya Member of the Villavieja Formation shows thin yellowish and reddish brown sandstone and siltstone levels with volcaniclastic grains.[20]
Honda area
The Honda Group extends for approximately 250 kilometres (160 mi) from the Upper to the Middle Magdalena Basin and is exposed in various locations along the right and left banks of the Magdalena River. Outcrops along the road between Girardot and Agua de Dios, Cundinamarca show a lower sequence of thick beds of greenish-yellow feldspar- and mica-rich conglomeratic sandstones, intercalated with reddish claystones and an upper level of alternating medium-to-coarse grained quartz arenites with low-angle cross stratification. These sandstones are intercalated with thick layers of fissile claystones with common calcareous sandstone concretions.[21] Sediments of the Honda Group restrict the course of the Sumapaz River to a narrow valley, close to its confluence with the Magdalena River.[22]
The majority of the municipality Prado, Tolima rests upon sediments of the Honda Group.[23] In the Middle Magdalena Basin and the eastern flank of the Central and the western flank of the Eastern Ranges, the group is subdivided into the Los Limones, San Antonio and Cambrás Formations. The total thickness of these formations in the northern original type section of the Honda Group reaches 3,255 metres (10,679 ft),[1] while a total thickness of 5,000 metres (16,000 ft) has been registered.[15]
Age
Thanks to the fossil abundance of the Honda Group at La Venta, the geological period of the sediments has received a separate name in the South American Mammal Ages (SALMA); Laventan, ranging from 13.8 to 11.8 Ma, as the only SALMA age defined north of the equator and in Colombia.[24] The age of the Villavieja Formation has been estimated to be between 17.0 and 12.1 Ma,[20] while the stratigraphically lower La Victoria Formation is dated at 13.82 to 12.38 Ma (Serravallian), based on fission track and volcanic analysis and paleomagnetic research.[18]
The Honda Group is laterally time equivalent with the lower part of the Real Formation in the central and northern Middle Magdalena Basin,[25] the lower part of the León Formation of the Llanos Basin,[26] the upper range of the Ciénaga de Oro Formation of the Lower Magdalena Basin,[27] and the Caja and Diablo Formations of the Llanos foothills.[28]
Depositional environment

The paleogeography of the meandering and braided river sediments of the Honda Group was characterised by a more African or Asian ecosystem than that of the Neotropics
The Honda Group has been deposited in a fluvial environment, with the lower part of the La Victoria Formation in a meandering setting, while the upper part was formed in a braided river system. The paleocurrent was from the west to the east and east-southeast.[19] The overlying predominantly finer grained Villavieja Formation was deposited in a meandering setting of a smaller size than those of the older La Victoria Formation. Paleocurrent analysis of the sediments in the Baraya and Cerro Colorado Members has revealed a similar flow direction as the La Victoria paleorivers, while the upper part of the Cerro Colorado Member shows an opposite trend to the west.[20] The volcanic clasts of the formations have as provenance the Central Range volcanism, of which the volcaniclastics in the Honda Group mark its onset.[29][30]
The depositional boundary for the Honda Group in the east is formed by the reverse Prado-Suárez and Cambrás-Salinas-Cambao Faults.[31]
Paleoclimate and vegetation
Analysis of the "Monkey Beds" of the Honda Group, provided estimates of annual precipitation levels between 1,500 and 2,000 millimetres (59 and 79 in). Today, these levels of rainfall are associated with the transition between savanna and forest environments in lowland South America. The vegetation of the La Venta fossil assemblage was diverse due to the different biomes of the depositional environment; meandering and braided river systems in a setting at lower altitudes than the present-day elevation of more than 400 metres (1,300 ft) above mean sea level. It has been suggested that the vegetational cover of the Honda Group sedimentary sequence was not a continuous canopy forest, yet a complex pattern of different flora ecosystems. The evergreen Amazonian foothill forests of today would therefore postdate the uplift of the Eastern Ranges of the Andes.[32] Based on vegetational and grazer diversity analysis of the La Venta fauna, it has been suggested the ecosystem resembled more that of Africa and Asia than of the modern Neotropics.[33] Research of the paleosols found in various levels within the Honda Group suggests the presence of arid areas in close proximity to pluvial parts.[34]
Petroleum geology
In the oil-producing Upper Magdalena Basin, the Honda Group is one of the reservoir formations, next to the more important Caballos and Monserrate Formations.[35][36] Shales of the Honda Group function as seal rock for certain oilfields in the Upper Magdalena Basin.[37] In the Tello Field in the basin, the Honda Group forms the overburden rock for the producing Monserrate reservoirs.[38]
Fossil content

A fossil species in the genus Arapaima has been described among the many fishes from the Honda Group
The Honda Group is the richest fossiliferous stratigraphic unit of Colombia, and one of the most important for the Miocene worldwide. At the La Venta site, numerous fossils of various orders have been recovered and are found still. The site marks a unique ecosystem showing a broad range of biodiversity. La Venta is also an important site as it represents the youngest uniquely South American faunal assemblage before the Great American Biotic Interchange; the result of the uplift of the Isthmus of Panama, of which the initial phase has been dated at around 12 Ma.[39] This led to a drastic alteration of the South American former island continental fauna. The Sparassodonta formed the dominant carnivorous mammal group in South America during most of the Cenozoic.
In South America, the carnivorous adaptive zone in terrestrial ecosystems was shared with other mammals; terror birds (Phorusrhacoidea), large crocodiles (Sebecidae), large snakes (Madsoiidae and Boidae), and even occasionally frogs.[40] The diversity of fossil freshwater fishes and crocodilians at La Venta is the richest assemblage of South America.[33][41] Fossilised trunks of Goupioxylon sp. have been identified in the Honda Group.[42]
The genus names Hondadelphys and Hondathentes, and the species epithets of Anadasypus hondanus and Scirrotherium hondaensis refer to the Honda Group. The giant sloth Brievabradys laventensis, the primate Stirtonia tatacoensis (originally described as Kondous laventicus), and the marsupial Micoureus laventicus were named after La Venta, while the primates Miocallicebus villaviejai and Stirtonia victoriae received their species epithets from the formations comprising the Honda Group.
Mammals
| Group | Fossils | Image | Notes |
|---|---|---|---|
| Primates | Aotus dindensis, Cebupithecia sarmientoi, Lagonimico conclucatus, Micodon kiotensis, Miocallicebus villaviejai, Mohanamico hershkovitzi, Neosaimiri annectens (syn. Laventiana annectens), Neosaimiri fieldsi, Nuciruptor rubricae, Patasola magdalenae, Stirtonia tatacoensis (syn. Kondous laventicus), Stirtonia victoriae |  | [43][44][45] [46][47][48] [49][50][51] [52][53][54] [55][56][57] |
| Astrapotheria | Hilarcotherium castanedaii |  | [58] |
| Granastrapotherium snorki |  | [59][60][61] | |
| Xenastrapotherium kraglievichi |  | [59] | |
| Anteaters and sloths | Hapalops sp. |  | [62] |
Brievabradys laventensis, Glossotheriopsis pascuali, Huilabradys magdaleniensis, Neonematherium flabellatum, Neotamandua borealis, Pseudoprepotherium confusum, Megalonychidae sp., Megatheriinae sp., Nothrotheriinae sp. | [63][62][64] [65][66] [67][68][69] | ||
| Armadillos | Anadasypus hondanus, Boreostemma acostae, B. gigantea, Nanoastegotherium prostatum, Neoglyptatelus originalis, Pedrolypeutes praecursor, Scirrotherium hondaensis, Dasypodidae sp. |  | [43][62][70] [71][72][73] [74][75] |
| Litopterna | Theosodon sp. |  | [59] |
Megadolodus molariformis, Pericotoxodon platignathus, Prolicaphrium sanalfolsensis, Prothoatherium colombianus, Villarroelia totoyoi, Proterotheriidae indet. | [59][76] [77][78][79] | ||
| Notoungulata | Huilatherium pluriplicatum |  | [80][59] |
Cochilius sp., Miocochilius anomopodus, Leontiniidae sp., Toxodontidae sp. | [59][81][82] | ||
| Sparassodonta | Anachlysictis gracilis |  | [43] |
| Dukecynus magnus |  | [43] | |
| Lycopsis longirostrus |  | [43][83] | |
| Hondadelphys fieldsi | [43][84][85] | ||
| Microbiotheria marsupials | Micoureus laventicus, Pachybiotherium minor | [43][86] | |
| Opossums | Thylamys colombianus, Thylamys minutus |  | [43][87][88] |
| Shrew opossums | Hondathentes cazador, Pithiculites chenche | [43][89][90] | |
| Rodents | Acarechimys minutissimus, Eodolichotis elachys, E. maddeni, Microscleromys cribiphilus, M. paradoxalis, Microsteiromys jacobsi, Neoreomys huilensis, Olenopsis sp., Prodolichotis guerreroi, P. pridiana, Rhodanodolichotis antepridiana, R. vucetichae, Ricardomys longidens, Scleromys colombianus, S. schurmanni, ?Steiromys sp., Dolichotinae sp., ?Echimyidae sp. | [44][62][91] [92][93][94] [95][96][97] [98][99] | |
| Bats | Diclidurus sp., Eumops sp., Kiotomops lopezi, Mormopterus colombiensis, Notonycteris magdalenensis, N. sucharadeus, Palynephyllum antimaster, Potamops mascahehenes, Tonatia sp. or Lophostoma sp. | [100][101] [102][103] | |
lesser bulldog bat (Noctilio albiventris), LaVal's disk-winged bat (Thyroptera lavali), Spix's disk-winged bat (Thyroptera tricolor), Thyroptera robusta | [100][104] | ||
| Sea cows | Potamosiren magdalenensis | [105] |
Birds
| Group | Fossils | Notes |
|---|---|---|
| Gruiformes | Aramus paludigrus | [106] |
| Piciformes | Galbula hylochoreutes | [106] |
| Opisthocomiformes | Hoazinoides magdalenae | [106][107] |
Reptiles and amphibians
| Group | Fossils | Image | Notes |
|---|---|---|---|
| Crocodiles | Gryposuchus colombianus |  | [108] |
| Langstonia huilensis |  | [109] | |
| Purussaurus neivensis |  | [110] | |
Balanerodus logimus, Charactosuchus fieldsi, Eocaiman sp., Gavialis sp., Mourasuchus atopus, Sebecus sp., Alligatoridae indet. | [61][111][112] [113][114] [115] | ||
| Turtles | Chelus colombiana, Geochelone hesterna, Podocnemis medemi | [116][117][118] | |
| Snakes | Colombophis portai, ?Eunectes stirtoni | [119][120] | |
| Lizards | Dracaena colombiana (syn. Paradracaena colombiana) | [121] | |
| Frogs | cane toad (Bufo marinus) | [122] |
Fish and crustaceans
| Group | Fossils | Notes |
|---|---|---|
| Fish | Colossoma macropomum, Lepidosiren paradoxa, cf. Acanthicus, Arapaima sp., Brachyplatystoma cf. B. vaillanti, Brachyplatystoma promagdalena, cf. Corydoras sp., Hoplias sp., cf. Hoplosternum, Hydrolycus sp., Phractocephalus hemiliopterus, Serrasalmus sp., Pygocentrus sp., or Pristobrycon sp. (cf. Myletes sp.), Ariidae gen. et. sp. Incertae sedis, Characidae cf. Tetragonopterinae gen. et. sp., Cichlidae gen. et. sp. Incertae sedis, Doradidae gen. et. sp. Incertae sedis 1-3, Loricariidae gen. et. sp. Incertae sedis 1 & 2, Potamotrygonidae | [41][123] [124][125] |
| Crabs | Sylviocarcinus piriformis | [41] |
Panorama

See also
- Geology of the Eastern Hills
- Geology of the Ocetá Páramo
- Geology of the Altiplano Cundiboyacense
Bogotá, Cerrejón, Floresta, Paja Formations
References
^ ab Acosta & Ulloa, 2002, p.34
^ Villarroel et al., 1996, p.47
^ ab Villarroel et al., 1996, p.48
^ abc Villarroel et al., 1996, p.50
^ ab Villarroel et al., 1996, p.51
^ López Quintero, 2007, p.1
^ Jiménez et al., 2012, p.186
^ Plancha 188, 2009
^ Kay & Madden, 1997, p.163
^ Sánchez et al., 2012, p.1
^ Parra, 2008, p.41
^ Villegas et al., 1994, p.1849
^ Carrera Ortíz, 2015, p.24
^ Álvarez et al., 2017, p.142
^ ab García et al., 2009. p.85
^ López Quintero, 2007, p.16
^ Villarroel et al., 1996, p.49
^ abc Villarroel, 1996, p.53
^ ab Villarroel et al., 1996, p.52
^ abcd Villarroel et al., 1996, p.54
^ ANi & Infracon, 2015, p.33
^ ANi & Infracon, 2015, p.45
^ N.N., s.a., p.182
^ Laventan at Fossilworks.org
^ Sánchez et al., 2012, p.3
^ García et al., 2009. p.60
^ Torres et al., 2012, p.2
^ Acosta & Ulloa, 2002, p.22
^ Gónzalez, 2001, p.81
^ Kroonenberg et al., 1982, p.21
^ Triviño, 1993, p.29
^ Kay & Madden, 1997, p.186
^ ab Kay & Madden, 1997, p.183
^ Flórez et al., 2013, p.243
^ Barrero et al., 2007, p.85
^ Carrera Ortíz, 2015, p.25
^ Carrera Ortíz, 2015, p.26
^ Ramón et al., 2001, p.63
^ De Porta, 2003, p.191
^ Súarez, 2015, p.03
^ abc (in Spanish) Fauna de La Venta - flora, cangrejos y peces
^ The Villavieja Formation, Colombia - Berkeley University
^ abcdefghi Croft, 2007, p.299
^ ab Croft, 2007, p.301
^ Meldrum & Kay, 1997
^ Organ & Lemelin, 2011
^ Rosenberger et al., 1991
^ Setoguchi & Rosenberger, 1988
^ Setoguchi, 1985
^ Takai et al., 2001
^ Tejedor, 2013, p.22
^ Defler, 2004, p.32
^ Cebupithecia sarmientoi at Fossilworks.org
^ Miocallicebus villaviejai at Fossilworks.org
^ Neosaimiri annectens at Fossilworks.org
^ Stirtonia tatacoensis at Fossilworks.org
^ Stirtonia victoriae at Fossilworks.org
^ Vallejo Pareja et al., 2015
^ abcdef Croft, 2007, p.302
^ Pardo Jaramillo, 2010
^ ab Granastrapotherium snorki, Sebecus sp. at Fossilworks.org
^ abcd Croft, 2007, p.300
^ Villarroel, 2000, p.118
^ Kay & Madden, 1997, p.171
^ Brievabradys laventensis at Fossilworks.org
^ Huilabradys magdaleniensis at Fossilworks.org
^ Neonematherium flabellatum at Fossilworks.org
^ Neotamandua borealis at Fossilworks.org
^ Pseudoprepotherium confusum at Fossilworks.org
^ Anadasypus hondanus at Fossilworks.org
^ Boreostemma acostae at Fossilworks.org
^ Boreostemma gigantea at Fossilworks.org
^ Neoglyptatelus originalis at Fossilworks.org
^ Pedrolypeutes praecursor at Fossilworks.org
^ Scirrotherium hondaensis at Fossilworks.org
^ Prolicaphrium sanalfolsensis at Fossilworks.org
^ Prothoatherium colombianus at Fossilworks.org
^ Duke Locality 33 at Fossilworks.org
^ Villarroelia totoyoi at Fossilworks.org
^ Villarroel, 1997
^ Villarroel et al., 1996, p.63
^ Hoffstetter, 1971, p.40
^ Suárez et al., 2015
^ Marshall, 1976
^ Hondadelphys fieldsi at Fossilworks.org
^ Pachybiotherium minor at Fossilworks.org
^ Thylamys colombianus at Fossilworks.org
^ Thylamys minutus at Fossilworks.org
^ Hondathentes cazador at Fossilworks.org
^ Pithiculites chenche at Fossilworks.org
^ Neoreomys huilensis at Fossilworks.org
^ Rhodanodolichotis antepridiana at Fossilworks.org
^ Scleromys colombianus at Fossilworks.org
^ Scleromys schurmanni at Fossilworks.org
^ Prodolichotis guerreroi at Fossilworks.org
^ UCMP V4936 at Fossilworks.org
^ Eodolichotis elachys at Fossilworks.org
^ Eodolichotis maddeni at Fossilworks.org
^ Rhodanodolichotis vucetichae at Fossilworks.org
^ ab Czaplewski et al., 2003, p.278
^ Takai et al., 1991
^ Kiotomops lopezi at Fossilworks.org
^ Potamops mascahehenes at Fossilworks.org
^ Thyroptera robusta at Fossilworks.org
^ Croft, 2007, p.303
^ abc Rasmussen, 1997
^ Miller, 1953
^ Gryposuchus colombianus at Fossilworks.org
^ Sebecus huiensis at Fossilworks.org
^ Purussaurus neivensis at Fossilworks.org
^ Charactosuchus fieldsi at Fossilworks.org
^ Mourasuchus atopus at Fossilworks.org
^ UCMP Locality V4524 at Fossilworks.org
^ Balanerodus logimus at Fossilworks.org
^ Eocaiman at Fossilworks.org
^ Cadena et al., 2008, p.1206
^ De la Fuente et al., 2013, p.102
^ Geochelone hesterna at Fossilworks.org
^ Hsiou et al., 2010
^ (in Spanish) Fauna de La Venta - serpientes
^ Dracaena colombiana at Fossilworks.org
^ Estes & Wassersug, 1963
^ Kay & Madden, 1997, p.185
^ Lundberg & Chernoff, 1992, p.5
^ Brachyplatystoma promagdalena at Fossilworks.org
Bibliography
Regional geology
Acosta Garay, Jorge E., and Carlos E. Ulloa Melo. 2002. Mapa Geológico del Departamento de Cundinamarca - 1:250,000 - Memoria explicativa, 1–108. INGEOMINAS. Accessed 2017-05-04.
Álvarez Rincón, Eduardo; Germán Rafael Bermúdez Arenas, and Lisette Daiyana Castillo Ramos. 2017. Elaboración del estudio de prefactibilidad para el montaje de una planta agroindustrial para la producción de carne de pescado en el municipio de Ricaurte, Cundinamarca, 1–264. Escuela Colombiana de Ingeniería Julio Garavito.
Barrero, Dario; Andrés Pardo; Carlos A. Vargas, and Juan F. Martínez. 2007. Colombian Sedimentary Basins: Nomenclature, Boundaries and Petroleum Geology, a New Proposal, 1–92. ANH.
Carrera Ortíz, Daniela. 2015. Sección balanceada del flanco occidental de la Cordillera Oriental en las inmediaciones de los municipios Villarrica y Cunday en la sub-cuenca de Girardot, Valle Superior del Magdalena, 1–68. EAFIT. Accessed 2017-05-18.
García González, Mario; Ricardo Mier Umaña; Luis Enrique Cruz Guevara, and Mauricio Vásquez. 2009. Informe Ejecutivo - evaluación del potencial hidrocarburífero de las cuencas colombianas, 1–219. Universidad Industrial de Santander.
González, Humberto. 2001. Geología de las planchas 206 Manizales y 225 Nevado del Ruíz - Memoria Explicativa, 1–93. INGEOMINAS. Accessed 2017-05-04.
Jiménez Díaz, Giovanny J.; Jhon Rico; Germán Bayona; Camilo Montes; Alexis Rosero, and Daniel Sierra. 2012. Analysis of curved folds and fault/fold terminations in the southern Upper Magdalena Valley of Colombia. Journal of South American Earth Sciences 39. 184–201.
Kroonenberg, Salomon B.; Hans Pichler, and Hans Diederix. 1982. Cenozoic alkalibasaltic to ultrabasic volcanism in the uppermost Magdalena Valley, Southern Huila Department, Colombia. Geología Norandina _. 19–26. Accessed 2017-05-04.
Parra, Mauricio. 2008. Cenozoic foreland-basin evolution in the northern Andes: insights from thermochronology and basin analysis in the Eastern Cordillera, Colombia (PhD thesis), 1–131. Universität Potsdam.
De Porta, Jaime. 2003. La formación del istmo de Panamá - su incidencia en Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales XXVII. 191–216.
Ramón, Juan Carlos; Gloria Vidal; Alexis Rosero; Pilar Gómez, and Hubert Borja. 2001. Revaluación del modelo geológico del Campo Tello (Valle Superior del Magdalena) y sus implicaciones en el programa de desarrollo. Geología Colombiana 26. 59–77. Accessed 2017-05-18.
Sánchez, Javier; Brian K. Horton; Eliseo Tesón; Andrés Mora; Richard A. Ketcham, and Daniel F. Stockli. 2012. Kinematic evolution of Andean fold-thrust structures along the boundary between the Eastern Cordillera and Middle Magdalena Valley basin, Colombia. Tectonics 31. 1–24. Accessed 2017-05-04.
Torres, Emilio J; Kurt J. Marfurt, and Carlos Molinares. 2012. Seismic characterization of delta front sand bodies in Lower Magdalena Valley basin and their potentiality as reservoirs in Cienaga de Oro Formation, Colombia, 1–5. SEG Las Vegas 2012 Annual Meeting.
Triviño Torres, Julio Alberto. 1993. Interpretación geotectónica e ingenieril en las subcuencas de Honda y Girardot, en imágenes de sensores remotos. Ingenería, Docencia e Investigación _. 24–36.
Villegas, M.E; Stefan Bachu; J.C. Ramon, and J.R. Underschultz. 1994. Flow of Formation Waters in the Cretaceous–Miocene Succession of the Llanos Basin, Colombia. AAPG Bulletin 78. 1843–1862.
N., N. s.a. Geología del Río Prado, 162–194. CORTOLIMA.
Local geology
ANi and, Infracon. 2015. Proyecto de asociación público de iniciativa privada ampliación tercer carril doble calzada Bogotá-Girardot, 1–231. Geotúneles Boma Inpasa. Accessed 2017-05-18.
Flórez M., María T.; Luis N. Parra S.; Daniel F. Jaramillo J., and José M. Jaramillo M. 2013. Paleosuelos del Mioceno en el Desierto de la Tatacoa. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales XXXVII. 229–244.
Kay, Richard F., and Richard H. Madden. 1997. Mammals and rainfall: paleoecology of the middle Miocene at La Venta (Colombia, South America). Journal of Human Evolution 32. 161–199. Accessed 2017-05-04.
López Quintero, Martha Isabel. 2007. Structural analysis of an area in the Neiva Sub-basin, Upper Magdalena Valley Basin, Colombia (MSc. thesis), 1–85. Colorado School of Mines.
Villarroel A., Carlos; Takeshi Setoguchi; Jorge Brieva, and Carlos Macia. 1996. Geology of the La Tatacoa "Desert" (Huila, Colombia): Precisions on the Stratigraphy of the Honda Group, the Evolution of the "Pata High" and the Presence of the La Venta Fauna. Memoirs of the Faculty of Science, Kyoto University. Series of geology and mineralogy 58. 41–66. Accessed 2017-05-04.
Paleontology
Cadena, Edwin; Carlos Jaramillo, and María E. Páramo. 2008. New material of Chelus colombiana (Testudines; Pleurodira) from the Lower Miocene of Colombia. Journal of Vertebrate Paleontology 28. 1206–1212. Accessed 2017-05-04.
Croft, Darin A. 2007. The Middle Miocene (Laventan) Quebrada Honda Fauna, southern Bolivia and a description of its Notoungulates. Palaeontology 50. 277–303. Accessed 2017-05-04.
Czaplewski, Nicolas J.; Masanaru Takai; Tiffany M. Naeher; Nobuo Shigehara, and Takeshi Setoguchi. 2003. Additional bats from the middle Miocene La Venta fauna of Colombia. Revista Académica Colombiana de Ciencias Naturales XXVII. 263–282. Accessed 2017-05-04.
Defler, Thomas. 2004. Historia natural de los primates colombianos, 1–613. Universidad Nacional de Colombia. Accessed 2017-05-04.
Estes, Richard, and Richard Wassersug. 1963. A Miocene toad from Colombia, South America. Breviora 193. 1–13. Accessed 2017-05-04.
De la Fuente, Marcelo; Juliana Sterli, and Ignacio Maniel. 2013. Origin, Evolution and Biogeographic History of South American Turtles, 1–168. Springer Science & Business Media. Accessed 2017-05-04.
Hoffstetter, Robert. 1971. Los vertebrados cenozóicos de Colombia: yacimientos, faunas, problemas planteados, 37–62. Universidad Nacional de Colombia. Accessed 2017-05-04.
Hsiou, Annie S.; Adriana M. Albino, and Jorge Ferigolo. 2010. Reappraisal of the south American Miocene snakes of the genus Colombophis, with description of a new species. Acta Palaeontologica Polonica 55. 365–379.
Lundberg, John G., and Barry Chernoff. 1992. A Miocene Fossil of the Amazonian Fish Arapaima (Teleostei, Arampaimidae) from the Magdalena River Region of Colombia - Biogeographic and Evolutionary Implications. Biotropica 24. 2–16. Accessed 2017-05-04.
Marshall, Larry G. 1976. New Didelphine Marsupials from the La Venta Fauna (Miocene) of Colombia, South America. Journal of Paleontology 50. 402–418. Accessed 2017-05-04.
Meldrum, D.J., and R.F. Kay. 1997. Nuciruptor rubricae, a new pitheciin seed predator from the Miocene of Colombia. American Journal of Physical Anthropology 102. 407–427. Accessed 2017-05-04.
Miller, Alden H. 1953. A fossil Hoatzin from the Miocene of Colombia. Auk 70. 484–495. Accessed 2017-05-04.
Organ, Jason M., and Pierre Lemelin. 2011. Tail Architecture and Function of Cebupithecia sarmientoi, a Middle Miocene Platyrrhine from La Venta, Colombia. The Anatomical Record 294.12. 2013–2023.
Pardo Jaramillo, Mauricio. 2010. Reporte de un nuevo ejemplar de Granastrapotherium snorki en el Valle Superior del Magdalena, Desierto de la Tatacoa, Huila. Colombia. Revista Académica Colombiana de Ciencias Naturales 34. 253–256. Accessed 2017-05-04.
Rasmussen, Tab. 1997. Vertebrate paleontology in the neotropics - the Miocene fauna of La Venta, Colombia, _. Smithsonian Institution Press.
Rosenberger, A.L.; T. Setoguchi, and W.C. Hartwig. 1991. Laventiana annectens, new genus and species: fossil evidence for the origins of callitrichine New World monkeys. Proceedings of the National Academy of Sciences of the United States of America 88. 2137–2140.
Setoguchi, Takeshi, and Alfred L. Rosenberger. 1988. A fossil owl monkey from La Venta, Colombia. Kyoto University overseas research reports of New World monkeys 6. 1–6. Accessed 2017-05-04.
Setoguchi, T. 1985. Kondous laventicus, a new ceboid primate from the Miocene of the La Venta, Colombia, South America. Folia Primatol 44. 96–101. Accessed 2017-05-04.
Suárez, Catalina; Analía M. Forasiepi; Franciso J. Goin, and Carlos Jaramillo. 2015. Insights into the Neotropics prior to the Great American Biotic Interchange: new evidence of mammalian predators from the Miocene of Northern Colombia. Journal of Vertebrate Paleontology e1029581. 1–10. Accessed 2017-05-04.
Takai, Masanaru; Federico Anaya; Hisashi Suzuki; Nobuo Shigehara, and Takeshi Setoguchi. 2001. A New Platyrrhine from the Middle Miocene of La Venta, Colombia, and the Phyletic Position of Callicebinae. Anthropological Science, Tokyo 109.4. 289.
Takai, Masanaru; Takeshi Setoguchi; Carlos Villarroel A.; Alberto Cadena G., and Nobuo Shigehara. 1991. A New Miocene Molossid Bat from La Venta, Colombia, South America. Memoirs of the Faculty of Science, Kyoto University. Series of geology and mineralogy 56. 1–9. Accessed 2017-05-04.
Tejedor, Marcelo F. 2013. Sistemática, evolución y paleobiogeografía de los primates Platyrrhini. Revista del Museo de La Plata 20. 20–39. Accessed 2017-05-04.
Vallejo Pareja, M.C.; J.D. Carrillo; J.W. Moreno Bernal; M. Pardo Jaramillo; D.F. Rodríguez González, and J. Muñoz Duran. 2015. Hilarcotherium castanedaii, gen. et sp. nov., a new Miocene astrapothere (Mammalia, Astrapotheriidae) from the Upper Magdalena Valley, Colombia. Zurich Open Repository and Archive, University of Zurich _. 1–14. Accessed 2017-05-04.
Villarroel A., Carlos S. 2000. Un nuevo Mylodontinae (Xenarthra, Tardigrada) en la Fauna de La Venta, Mioceno de Colombia: el estado actual de la Familia Orophodontidae. Revista de la Academia Colombiana de Ciencias Exactas Físicas y Naturales 24. 117–127. Accessed 2017-05-04.
Villarroel A., Carlos S. 1997. La Estructura de la Dentición Caduca de Huilatherium pluriplicatum, Leontiniidae (Notoungulata) del Mioceno de Colombia. Geología Colombiana 22. 139–149. Accessed 2017-05-04.
Wheeler, Brandon. 2010. Community ecology of the Middle Miocene primates of La Venta, Colombia: the relationship between ecological diversity, divergence time, and phylogenetic richness. Primates 51.2. 131–138.
Maps
Barrero L., Darío, and Carlos J. Vesga O. 2009. Plancha 188 - La Dorada - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
Barrero, Darío, and Carlos J. Vesga. 2010. Plancha 207 - Honda - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
Barrero, Darío, and Carlos J. Vesga. 2010. Plancha 226 - Líbano - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
Acosta, Jorge E.; Rafael Guatame; Oscar Torres, and Frank Solano. 1999. Plancha 245 - Girardot - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
Nuñez, Alberto; Dario Mosquera, and Carlos J. Vesga. 2009. Plancha 263 - Ortega - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
Calcedo, Juan Carlos, and Roberto Terraza. 2000. Plancha 264 - Espinal - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
Carvajal, Cesar; Jaime Fuquen, and Luis Gómez. 1993. Plancha 282 - Chaparral - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
Cossio, Ubaldo; Gabriel Rodríguez, and Miguel Rodríguez. 1995. Plancha 283 - Purificación - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
Fuquen, Jaime; Gabriel Rodríguez; Ubaldo Cossio, and Alberto Núñez. 1993. Plancha 302 - Aipe - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
Acosta, Jorge; Pablo Caro; Jaime Fuquen, and José Osorno. 2002. Plancha 303 - Colombia - 1:100,000, 1. INGEOMINAS.
Ferreira, Paulina; Alberto Núñez, and Miguel Rodríguez. 1998. Plancha 323 - Neiva - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
Marquínez, Germán; C. Morales, and Alberto Núñez. 1999. Plancha 344 - Tesalia - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
Velandia, F.; C.J. Morales; J.C. Caicedo, and Alberto Núñez. 1999. Plancha 345 - Campoalegre - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
Rodríguez, Gabriel; Paulina Ferreira; Francisco Velandia, and Alberto Núñez. 1998. Plancha 366 - Garzón - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
Rodríguez, Gabriel; Gilberto Zapata, and M. Velázquez. 2003. Plancha 367 - Gigante - 1:100,000, 1. INGEOMINAS. Accessed 2017-06-06.
External links
| Wikimedia Commons has media related to Honda Group, Colombia. |
(in Spanish) Faunas del Cenozoico de Colombia
(in Spanish) Fauna de La Venta - aspectos geológicos
South American land mammal ages | |||||||
|---|---|---|---|---|---|---|---|
| Ages |
|  | |||||
| Defining formations and sites |
| ||||||
| Sources |
| ||||||
Paleontology in Colombia | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Notable researchers |
|     | |||||||||||||||||||||||||
| Major fossiliferous stratigraphic units |
| ||||||||||||||||||||||||||
| Notable fossils |
| ||||||||||||||||||||||||||


