Crown group
Two distinct crown groups (in red) are illustrated, connected by an ancestor (black circle). The two groups form a larger crown group (lilac).
| Part of a series on |
| Evolutionary biology |
|---|
Key topics
|
Processes and outcomes
|
Natural history
|
History of evolutionary theory
|
Fields and applications
|
Social implications
|
|
In phylogenetics, the crown group of a collection of species consists of the living representatives of the collection together with their ancestors back to their most recent common ancestor as well as all of that ancestor's descendants. It is thus a clade, a group consisting of a species and all its descendants.
The concept was developed by Willi Hennig, the formulator of phylogenetic systematics, as a way of classifying living organisms relative to their extinct relatives in his "Die Stammesgeschichte der Insekten",[1]
and the "crown" and "stem" group terminology was coined by R. P. S. Jefferies in 1979.[2] Though formulated in the 1970s, the term was not commonly used until its reintroduction in 2000 by Graham Budd and Sören Jensen.[3]
Contents
1 Contents of the crown group
2 Other groups under the crown group concept
2.1 Pan-group
2.2 Stem groups
2.2.1 Examples of stem groups (in the wider sense)
2.3 Plesion-group
3 Palaeontological significance of stem and crown groups
4 Stem groups in systematics
5 References
6 Further reading
Contents of the crown group
It is not necessary for a species to have living descendants in order for it to be included in the crown group. Extinct side branches on the family tree that are descended from the most recent common ancestor of living members will still be part of a crown group. For example, if we consider the crown-birds (i.e. all extant birds and the rest of the family tree back to their most recent common ancestor), extinct side branches like the dodo or great auk are still descended from the most recent common ancestor of all living birds, so fall within the bird crown group.[4] One very simplified cladogram for birds is shown below:[5]
.mw-parser-output table.clade{border-spacing:0;margin:0;font-size:100%;line-height:100%;border-collapse:separate;width:auto}.mw-parser-output table.clade table.clade{width:100%}.mw-parser-output table.clade td{border:0;padding:0;vertical-align:middle;text-align:center}.mw-parser-output table.clade td.clade-label{width:0.8em;border:0;padding:0 0.2em;vertical-align:bottom;text-align:center}.mw-parser-output table.clade td.clade-slabel{border:0;padding:0 0.2em;vertical-align:top;text-align:center}.mw-parser-output table.clade td.clade-bar{vertical-align:middle;text-align:left;padding:0 0.5em}.mw-parser-output table.clade td.clade-leaf{border:0;padding:0;text-align:left;vertical-align:middle}.mw-parser-output table.clade td.clade-leafR{border:0;padding:0;text-align:right}
Aves |
| ||||||||||||
| |
In this diagram, the clade labelled "Neornithes" is the crown group of birds: it includes the most recent common ancestor of all living birds and its descendants, living or not. Although considered to be birds (i.e. members of the clade Aves), Archaeopteryx and other extinct groups are not included in the crown group, as they fall outside the Neornithes clade, being descended from an earlier ancestor.
An alternative definition does not require any members of a crown group to be extant, only to have resulted from a "major cladogenesis event".[6] The first definition forms the basis of this article.
Often, the crown group is given the designation "crown-", to separate it from the group as commonly defined. Both birds and mammals are traditionally defined by their traits,[7][8] and contain fossil members that lived before the last common ancestors of the living groups or, like the mammal Haldanodon,[9] were not descended from that ancestor although they lived later. Crown-Aves and Crown-Mammalia therefore differ slightly in content from the common definition of Aves and Mammalia. This has caused some confusion in the literature.[10][11]
Other groups under the crown group concept
The cladistic idea of strictly using the topology of the phylogenetic tree to define groups, necessitates other definitions than crown groups to adequately define commonly discussed fossil groups like various Burgess Shale fauna. Thus, a host of prefixes have been defined to describe various branches of the phylogenetic tree relative to extant organisms.[12]
Pan-group
A pan-group or total group is the crown group and all organisms more closely related to it than to any other extant organisms. In a tree analogy, it is the crown group and all branches back to (but not including) the split with the closest branch to have living members. The Pan-Aves thus contain the living birds and all (fossil) organisms more closely related to birds than to crocodilians (their closest living relatives). The phylogenetic lineage leading back from Neornithes to the point where it merges with the crocodilian lineage, along with all side branches, constitutes pan-birds. In addition to non-crown group primitive birds like Archaeopteryx, Hesperornis and Confuciusornis, therefore, pan-group birds would include all dinosaurs and pterosaurs as well as an assortment of non-crocodilian animals like Marasuchus.
Pan-Mammalia consists of all mammals and their fossil ancestors back to the phylogenetic split from the remaining amniotes (the Sauropsida). Pan-Mammalia is thus an alternative name for Synapsida.
Stem groups
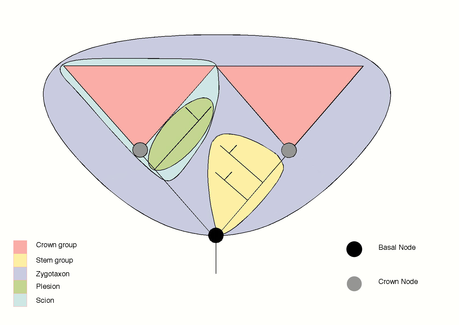
A stem group is a paraphyletic group composed of a pan-group or total group, above, minus the crown group itself (and therefore minus all living members of the pan-group). This leaves primitive relatives of the crown groups, back along the phylogenetic line to (but not including) the last common ancestor of the crown group and their nearest living relatives. It follows from the definition that all members of a stem group are extinct. The "stem group" is the most used and most important of the concepts linked to crown groups, as it offers a purely phylogenetic route to classify fossils that otherwise do not fit into systematics based on living organisms.
While often attributed to Jefferies (1979), Willmann (2003)[13] traced the origin of the stem group concept to German systematist Othniel Abel (1914)
,[14] and it was discussed and diagrammed in English as early as 1933 by A. S. Romer.[15]
Alternatively, the term "stem group" is sometimes used in a narrower sense to cover just the members of the traditional taxon falling outside the crown group. Permian synapsids like Dimetrodon and Anteosaurus are stem mammals in the wider sense but not in the narrower one.[16]
Examples of stem groups (in the wider sense)
Stem birds perhaps constitute the most cited example of a stem group, as the phylogeny of this group is fairly well known. The following cladogram, based on Benton (2005),[17] illustrates the concept:
| stem group birds crown group birds |
The crown group here is Neornithes, all modern bird lineages back to their last common ancestor. The closest living relatives of birds are crocodilians. If we follow the phylogenetic lineage leading to Neornithes to the left, the line itself and all side branches belong to the stem birds until the lineage merges with that of the crocodilians. In addition to non-crown group primitive birds like Archaeopteryx, Hesperornis and Confuciusornis, stem group birds include the dinosaurs and the pterosaurs. The last common ancestor of birds and crocodilians—the first crown group archosaur—was neither bird nor crocodilian and possessed none of the features unique to either. As the bird stem group evolved, distinctive bird features such as feathers and hollow bones appeared. Finally, at the base of the crown group, all traits common to extant birds were present.
Stem mammals are those in the lineage leading to living mammals, together with side branches, from the divergence of the lineage from the Sauropsida to the last common ancestor of the living mammals. This group includes the synapsids as well as mammaliaforms like the morganucodonts and the docodonts; the latter groups are traditionally and anatomically been considered mammals even though they fall outside the crown group mammals.[18]
Stem tetrapods are the animals belonging to the lineage leading to tetrapods from their divergence from the lungfish, our nearest relatives among the fishes. In addition to a series of lobe-finned fishes, they also include some of the early labyrinthodonts. Exactly what labyrinthodonts are in the stem group tetrapods rather than the corresponding crown group is uncertain, as the phylogeny of early tetrapods is not well understood.[19] This example shows that crown and stem group definitions are of limited value when there is no consensus phylogeny.
Stem arthropods constitute a group that has seen attention in connection with the Burgess Shale fauna. Several of the finds, including the enigmatic Opabinia and Anomalocaris have some, though not all, features associated with arthropods, and are thus considered stem arthropods.[20][21] The sorting of the Burgess Shale fauna into various stem groups finally enabled phylogenetic sorting of this enigmatic assemblage and also allowed for identifying velvet worms as the closest living relatives of arthropods.[21]
Stem priapulids are other early Cambrian to middle Cambrian faunas, appearing in Chengjiang to Burgess Shale. The genus Ottoia has more or less the same build as modern priapulids, but phylogenetic analysis indicates that it falls outside the crown group, making it a stem priapulid.[3]
Plesion-group
The name plesion has a long history in biological systematics, and plesion group has acquired several meanings over the years. One use is as "nearby group" (plesion means close to in Greek), i.e. sister group to a given taxon, whether that group is a crown group or not.[22] The term may also mean a group, possibly paraphyletic, defined by primitive traits (i.e. symplesiomorphies).[23] It is generally taken to mean a side branch splitting off earlier on the phylogenetic tree than the group in question.
Palaeontological significance of stem and crown groups
Placing fossils in their right order in a stem group allows the order of these acquisitions to be established, and thus the ecological and functional setting of the evolution of the major features of the group in question. Stem groups thus offer a route to integrate unique palaeontological data into questions of the evolution of living organisms. Furthermore, they show that fossils that were considered to lie in their own separate group because they did not show all the diagnostic features of a living clade, can nevertheless be related to it by lying in its stem group. Such fossils have been of particular importance in considering the origins of the tetrapods, mammals, and animals.
The application of the stem group concept also influenced the interpretation of the organisms of the Burgess shale. Their classification in stem groups to extant phyla, rather than in phyla of their own, is thought by some to make the Cambrian explosion easier to understand without invoking unusual evolutionary mechanisms;[24] however, application of the stem group concept does nothing to ameliorate the difficulties that phylogenetic telescoping[25][26] poses to evolutionary theorists attempting to understand both macroevolutionary change and the abrupt character of the Cambrian explosion. Overemphasis on the stem group concept threatens to delay or obscure proper recognition of new higher taxa.[27]
Stem groups in systematics
As originally proposed by Karl-Ernst Lauterbach, stem groups should be given the prefix "stem" (i.e. Stem-Aves, Stem-Arthropoda), however the crown group should have no prefix.[28] The latter has not been universally accepted for known groups. A number of paleontologists have opted to apply this approach anyway.[29]
References
^ Hennig, W. (1969), Die Stammesgeschichte der Insekten, Waldemar Kramer, Frankfurt am Main.mw-parser-output cite.citation{font-style:inherit}.mw-parser-output q{quotes:"""""""'""'"}.mw-parser-output code.cs1-code{color:inherit;background:inherit;border:inherit;padding:inherit}.mw-parser-output .cs1-lock-free a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-limited a,.mw-parser-output .cs1-lock-registration a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-subscription a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration{color:#555}.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration span{border-bottom:1px dotted;cursor:help}.mw-parser-output .cs1-hidden-error{display:none;font-size:100%}.mw-parser-output .cs1-visible-error{font-size:100%}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-format{font-size:95%}.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-left{padding-left:0.2em}.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-right{padding-right:0.2em}
^ Jefferies, R.P.S. (1979). "The Origin of Chordates — A Methodological Essay". In M. R. House. The Origin of Major Invertebrate Groups. London ; New York: Academic Press for The Systematics Association. pp. 443–447. ISBN 0123574501.
^ ab Budd, G.E.; Jensen, S. (2000), "A critical reappraisal of the fossil record of the bilaterian phyla", Biological Reviews, 75 (2): 253–295, doi:10.1111/j.1469-185X.1999.tb00046.x, PMID 10881389
^ "DNA yields dodo family secrets". BBC News. London. 2002-02-28. Retrieved 2006-09-07.
^ Chiappe, Luis M. (2007), Glorified Dinosaurs: The Origin and Early Evolution of Birds, Sydney: University of New South Wales Press, ISBN 978-0-86840-413-4
^ UCMP Glossary: Phylogenetics, University of California Museum of Paleontology, retrieved 2010-12-04
^ ed, Rob Nagel, (2001). U-X-L encyclopedia of science (2nd ed.). Detroit: U-X-L. ISBN 0787654329.
^ Benton, Michael J. (2004). Vertebrate palaeontology (3rd ed.). Oxford: Blackwell Science. ISBN 0632056371.
^ Luo, Zhe-Xi (2007). "Transformation and diversification in early mammal evolution" (PDF). Nature. 450 (7172): 1011–1019. Bibcode:2007Natur.450.1011L. doi:10.1038/nature06277. PMID 18075580. Archived from the original (PDF) on 2012-11-24.
^ Anderson, Jason S. (2002). "Use of Well-Known Names in Phylogenetic Nomenclature: A Reply to Laurin" (PDF). Systematic Biology. 51 (5): 822–827. doi:10.1080/10635150290102447. PMID 12396594. Retrieved 28 December 2011.
^ Laurin, M.; Anderson, J.S. (2004). "Meaning of the Name Tetrapoda in the Scientific Literature: An Exchange". Systematic Biology. 53: 68–80. doi:10.1080/10635150490264716. PMID 14965901.
^ Craske, A. J. & Jefferies, R. P. S. (1989): A new mitrate from the Upper Ordovician of Norway, and a new approach to subdividing a plesion. Palaeontology no 32, pages 69–99
^ Willmann, Rainer (2003). "From Haeckel to Hennig: the early development of phylogenetics in German-speaking Europe". Cladistics. 19: 449–479. doi:10.1016/j.cladistics.2003.09.001.
^ Abel, O. (1914), Die vorzeitlichen Saugetiere, G. Fischer Verlag, Jena
^ Romer, A.S. (1933), Vertebrate Paleontology, University of Chicago Press, Chicago
^ Czaplewski, Terry A. Vaughan, James M. Ryan, Nicholas J. (2000). Mammalogy (4th ed.). Fort Worth: Brooks/Cole Thomson Learning. p. 61. ISBN 003025034X. Retrieved 27 July 2013.From a cynodont ancestry, the stem mammals arose in the late Triassic, slightly after the first appearance of dinosaurs
^ Benton, M. J. (2005). Vertebrate Palaeontology (3rd ed.). Blackwell Publishing. ISBN 978-0-632-05637-8.
^ Romer, A.S. (1966): Vertebrate Paleontology. University of Chicago Press, Chicago; 3rd edition
ISBN 0-7167-1822-7
^ Marjanović, David; Laurin, Michel (1 March 2013). "The origin(s) of extant amphibians: a review with emphasis on the "lepospondyl hypothesis"". Geodiversitas. 35 (1): 207–272. doi:10.5252/g2013n1a8.
^ Budd, G.E. (1996), "The morphology of Opabinia regalis and the reconstruction of the arthropod stem-group", Lethaia, 29 (1): 1–14, doi:10.1111/j.1502-3931.1996.tb01831.x.
^ ab Brysse, K. (2008), "From weird wonders to stem lineages: the second reclassification of the Burgess Shale fauna", Studies in History and Philosophy of Science Part C: Biological and Biomedical Sciences, 39 (3): 298–313, doi:10.1016/j.shpsc.2008.06.004, PMID 18761282.
^ Patterson, C.; Rosen, D.E. (1977). "Review of ichthyodectiform and other Mesozoic teleost fishes and the theory and practice of classifying fossils". Bulletine of American Natural History. 158 (2): 85–172.
^ Kluge, N. (2000). Modern Systematics of Insects. Part I. Principles of Systematics of Living Organisms and General System of Insects, with Classification of Primary Wingless and Paleopterous Insects (in Russian). St. Petersburg, Russland: Lan'. p. 336.
^ Brysse, K. (2008), "From weird wonders to stem lineages: the second reclassification of the Burgess Shale fauna", Studies in History and Philosophy of Science Part C, 39 (3): 298–313, doi:10.1016/j.shpsc.2008.06.004, PMID 18761282
^ Conway Morris, S. (2009). "Walcott, the Burgess Shale, an rumours of a post-Darwinian world". Current Biology. 19 (20): R927–R931. doi:10.1016/j.cub.2009.08.046. PMID 19889363.
^ McMenamin, M. A. S. (2013). "Breakthrough on the Cambrian Explosion". BioScience. 63 (10): 834–835. doi:10.1525/bio.2013.63.10.14.
^ McMenamin, M. A. S. (2015). Paramphibia: A New Class of Tetrapods. South Hadley, Massachusetts: Meanma. doi:10.13140/2.1.2569.0401. ISBN 978-1-893882-20-1.
^ Lauterbach, K-E. (1989): Das Pan-Monophylum – ein Hilfsmittel für die Praxis der Phylogenetischen Systematik. Zoologischer Anzeiger, no 223, pp 139–156.
^ Gauthier, J., and de Queiroz, K. (2001). "Feathered dinosaurs, flying dinosaurs, crown dinosaurs, and the name Aves." Pp. 7-41 in New perspectives on the origin and early evolution of birds: proceedings of the International Symposium in Honor of John H. Ostrom (J. A. Gauthier and L. F. Gall, eds.). Peabody Museum of Natural History, Yale University, New Haven, Connecticut, U.S.A.
Further reading
Budd, G. (2001), "Climbing Life's Tree", Nature, 412 (6846): 487, doi:10.1038/35087679, PMID 11484029
Budd, G. E. (2001), "Tardigrades as 'stem-group' Arthropods: the Evidence from the Cambrian Fauna", Zoologischer Anzeiger, 240 (3–4): 265–279, doi:10.1078/0044-5231-00034
Craske, A. J.; Jefferies, R. P. S. (1989), "A New Mitrate from the Upper Ordovician of Norway, and a New Approach to Subdividing a Plesion", Palaeontology, 32: 69–99
Phylogenetics | |
|---|---|
| Relevant fields |
|
| Basic concepts |
|
| Inference methods |
|
| Current topics |
|
| Group traits |
|
| Group types |
|
| Nomenclature |
|
| |


